
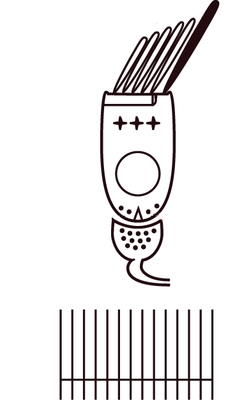
The semicircular canals consist of membranous ducts where angular/rotational acceleration and velocity of the head are sensed. As the semicircular canal ends, the duct enlarges to form the ampulla.
The semicircular canal system functions as a three-dimensional, bi-directional angular accelerometer for head movements1.
The semicircular canals are stimulated by angular acceleration, and the neural firing rate depends on the magnitude of head angular acceleration. However, the neural output from the sensory cells represents the velocity of rotation. This mathematical integration of the input signal is due to the mechanics of the canals, mainly the increase in viscous properties of the fluid due to the small size of the canal, relative to its radius of curvature2.
The directional sensitivity of the semicircular canals is obtained by resolving head movement into three vectors. All the hair cell receptors in the crista of each canal are aligned along the axis of their respective canal and the hair cells in each crista are oriented with their kinocilia in the same direction, so that the cells all have the same directional sensitivity.
The anatomic orientation of the semicircular canals therefore determines the direction of these three vectors of head movement3 (this is completely different to the otoliths of the utricle and saccule, which have hair cells aligned in a multitude of directions).
For rotational motion, vestibular afferents are able to discriminate differences in head velocity of the order of 2 deg/sec, but they are also sensitive to a broad range of natural head movements up to high head speeds of hundreds of deg/sec (as one might experience when making a fast head turn toward a loud sound, or performing gymnastics4.

Redrawn from:The Human Memory. Vestibular System. Retrieved from https://human-memory.net/vestibular-system/
RA, right anterior RP, right posterior RH, right horizontal
LA, left anterior LP, left posterior LH, left horizontal

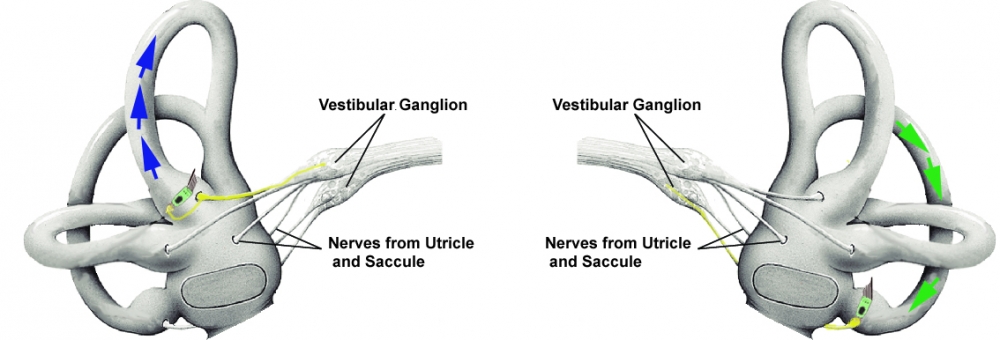
Pairing of semi-circular canals allows for redundant reception of movement, explaining compensation after unilateral vestibular loss:
Pairs of canals detect head motion in the same plane:
- Right Lateral-Left Lateral
- Right Anterior-Left Posterior
- Left Anterior-Right Posterior

The anterior and posterior canals are oriented vertically at 45 degrees to the sagittal axis, so that both canals respond to pitch and roll movements.
The horizontal canals respond maximally to yaw. The mirror symmetry of the canals across the head means that the two horizontal canals provide similar, although inverse signals. Working like a push–pull mechanism, one canal in the pair elicits excitation, whereas the other canal causes inhibition (depending on the direction of rotation). Corresponding to this arrangement, there are cells in the vestibular nuclei that are excited by a particular canal but inhibited by its opposite number on the other side, thus effectively combining the signals from the two sides in a push–pull manner.
- Yaw rotation of the head to the right increases the firing of afferents from the right horizontal canal but decreases the firing rate from the left horizontal canal. The right horizontal semicircular canal stimulates right CN 8 discharge in response to rightward head rotation (and inhibits right CN 8 discharge in response to leftward head rotation). Conversely, the left horizontal semicircular canal excites the left CN 8 in response to leftward head rotation and inhibits the left CN 8 in response to rightward head rotation.
- The other canals operate similarly to the horizontal canal, with each anterior canal being aligned with the contralateral posterior canal6. For example, when bending forward and to the side, one anterior canal is stimulated and the posterior canal in the opposite labyrinth is inhibited6. The anterior semicircular canals stimulate upgaze (in response to downward head rotation), and the posterior canals stimulate downgaze (in response to upward head rotation). The anterior and posterior canals also stimulate contralateral torsion in response to dynamic ipsilateral head tilt.
Each of the three semicircular canals defines a plane; head rotation about an axis perpendicular to this plane is the most effective stimulus, whereas rotations about axes lying in this plane are ineffective6.


(vv)Equilibrium The Ampulla In The Posterior Semicircular Canal.mp4(tt)

Cristae: these organs lie within the ampulla of the semi-circular canals; the crista crosses each ampulla in a direction perpendicular to the longitudinal axis of the canal.
Long stereocilia of hair cells are located on the surface of the crista, with the cilia protruding into and embedded in the cupula, a gelatinous mass that extends from the surface of the crista to the ceiling of the ampulla, forming a watertight seal.
Head motion is detected because the semicircular canals are filled with endolymph. When the head begins to rotate, the bony canals move with it. However, the viscous endolymph is not physically attached to the head, and therefore lags behind due to inertia. Thus, relative to the canals, the endolymph rotates in the opposite direction of head turn. The rotating endolymph displaces the cupula and stimulates the stereocilia, causing a neural signal which indicates the rotation6. With the three semicircular canals aligned at right angles to each other, rotation in any direction can be resolved.
Since the cupula has the same specific gravity as the surrounding endolymph, it is not subject to gravitational forces. The motion of the cupula can be likened to that of a pendulum in a viscous medium; sudden displacement of the cupula by an abrupt change in angular velocity is followed by a gradual exponential return of the cupula to its baseline position.
During sinusoidal head rotation in the plane of a semi-circular canal, a sinusoidal change in firing frequency is superimposed on the high baseline resting discharge of the canal. The peak firing rate occurs at the time of maximum cupular displacement, which occurs at the time of peak angular head velocity. For large amplitude sinusoidal rotations, the responses of the horizontal semicircular canals becomes increasingly asymmetrical. The excitatory responses can increase to more than 400 spikes per second in proportion to stimulus magnitude, whereas the growth of inhibitory response is limited to the disappearance of spontaneous activity. This asymmetry in afferent nerve response partially explains the presence of a positive head thrust sign in patients with only one functioning labyrinth.
With rotation of the head, the slow phase of nystagmus and endolymph move in the same direction: a spin to the right results in right beating nystagmus since the endolymph moves to the left due to inertia (opposite to the direction of head movement), with the slow phase of nystagmus to the left, and the quick phase to the right.
(vv)Endolymph Motion.mp4(tt)
The basic element of the labyrinthine receptor organs that transduces mechanical force to nerve action potentials is the hair cell.
A bundle of nonmobile stereocilia protrudes from the apical end of each hair cell. The height of the stereocilia increases stepwise from one side to the other, and next to the tallest stereocilium is a thicker, longer cilia, the kinocilium. The tips of the cilia are connected by tip-links that open and close mechanosensory channels8.
Each afferent neuron has a baseline firing rate:
- Deflection of stereocilia towards the kinocilium results in an increase in the firing rate of the afferent neuron in the horizontal canal: this opens the mechanosensory channels at the tips causing an influx of potassium and depolarization of the resting membrane potential, and opens voltage-gated calcium channels at the base, with release of neurotransmitter (mostly glutamate) activating the afferent nerve terminals.
- Deflection away from the kinocilium causes a decrease in the firing rate in the horizontal canal: closing of the channels and hyperpolarization of the hair cells.
There is continuous spontaneous activity of the afferent nerves of the hair cells: a small percentage of the mechano-sensory channels remains open at rest, leading to spontaneous afferent nerve discharges. The spontaneous firing rate varies among different animal species and among different sensory receptors, and is thought to be highest in the afferent neurons of the semicircular canals of mammals (up to 100 spikes per second).
The forces associated with angular head acceleration displace the cupula and bend the hair cells of the crista:

Redrawn from: Baloh RW, Kerber, K. Baloh and Honrubia's Clinical Neurophysiology of the Vestibular System. Oxford University Press; 2010.
In the horizontal canal, the kinocilia are directed toward the utricular side of the ampulla, whereas in the vertical canals the kinocilia are directed toward the canal side of the ampulla. This explains the difference in directional sensitivity between horizontal and vertical canals:
- the afferent nerve fibers of the horizontal canals increase their baseline firing when endolymph moves toward the utricle and ampulla (ampullopetal flow)
- the afferent nerves of the vertical canals increase their baseline firing rate with endolymph flow away from the ampulla (ampullofugal flow)
When the head turns to the left, the cupula is pushed toward the kinocilium in the left horizontal canal, and the firing rate of the relevant axons in the left vestibular nerve increases. Conversely, the cupula in the right horizontal canal is pushed away from the kinocilium, with a concomitant decrease in the firing rate of the related neurons.

In the vertical canals, kinocilia are oriented towards canal side of ampulla
![]()
Arrow shows direction of the endolymph movement, which is towards the cupula (ampullopetal) and excitatory: there is a resultant displacement of the stereocilia and increased discharge in the vestibular nerve of the lateral canal. (Note that the kinocilia of the horizontal canal are orientated towards the utricular side of the canal).
Right vestibular apparatus viewed from the front
Arrows shows direction of the endolymph movement, and resultant displacement of the stereocilia to activate the right superior canal, and to inhibit the left posterior canal.
Note that the kinocilia of the superior and posterior canals are orientated towards the canal side of their respective ampulla.
Redrawn from:T. G. Constandinou, J. Georgiou and C. Toumazou, "A neural implant ASIC for the restoration of balance in individuals with vestibular dysfunction," 2009 IEEE International Symposium on Circuits and Systems, Taipei, 2009, pp. 641-644, doi: 10.1109/ISCAS.2009.5117830
Endolymph moves in the opposite direction to head rotation, deflecting the cupula away from the canal with an excitatory ampullopetal stimulus.
In the case of head turning to the left, the endolymph moves in the opposite direction, an ampullofugal stimulus, and the resulting displacement of the cupula increases the resting discharge from the left horizontal canal.

Redrawn from: Angelaki D, Dickman JD. The Vestibular System.. Retrieved from https://nobaproject.com/modules/the-vestibular-system#content
If the head rotates to the left, the left horizontal canal will be excited, and the right inhibited..
In order to keep the eyes fixed on a stationary point, the right lateral rectus and left medial rectus must fire, to contract these muscle and move the eyes to the right.
The pathway is as follows:
- Head rotation to the left causes endolymph to flow in the opposite direction (right), this stimulates the hair cells in the cupula, exciting the left horizontal canal.
- The vestibular nerve picks up the signals coming from the hair cells, enters the brainstem and synapses in the vestibular nucleus.
- Fibers from the vestibular nucleus cross over to the contralateral side and project to the abducens nucleus (VI) on the right side, to stimulate the contralateral (right) lateral rectus.
- They also project to the occulomotor nucleus (III) on the ipsilateral side, to stimulate the ipsilateral (left) medial rectus.
- The same vestibular cells also inhibit the opposing muscles (in this case, the right medial rectus, and the left lateral rectus).
On the other side, the right horizontal canal is wired to the complementary set of muscles. Since it is inhibited, it will not excite its target muscles (the right medial rectus and the left lateral rectus), nor will it inhibit the muscles moving the eyes to the right (the right lateral rectus and the left medial rectus).