
The saccadic command system depends on two major entities, the superior colliculus, and cortical regions responsible for saccades, such as the frontal eye field, parietal eye field and supplementary eye field.
Superior Colliculus
The superior colliculus (SC) contains a motor map that reflects direction and size of saccades.
- The rostral SC is sensory and reflects the foveal region of the retina, and receives information from the lateral geniculate body and visual cortex. The cells are arranged in an
orderly way on the surface of the colliculus, forming a map of visual space.
This region has neurons that discharge continuously during visual fixation, and stimulating rostral SC suppresses the generation of saccadic eye movements by stimulating the omnipause neurons1. - The caudal SC is essentially motor: it projects to brainstem and evokes saccades directed to the contralateral side; this caudal “saccade zone” of the SC triggers saccades and drives the excitatory and inhibitory burst neurons1,2. The caudal SC is silent during fixation. Just before the onset of a saccade, the caudal SC becomes active, and the rostral SC shuts off. Appearance of a target will activate cells in the caudal SC at a site corresponding to the target’s retinal location, and electrical stimulation of the caudal SC at that same locus will evoke a saccade to that target location1.
Ablations of the SC significantly increase saccade latency and reduce peak speed, but do not affect accuracy.
Vertical broken lines indicate onset and end of horizontal saccade.
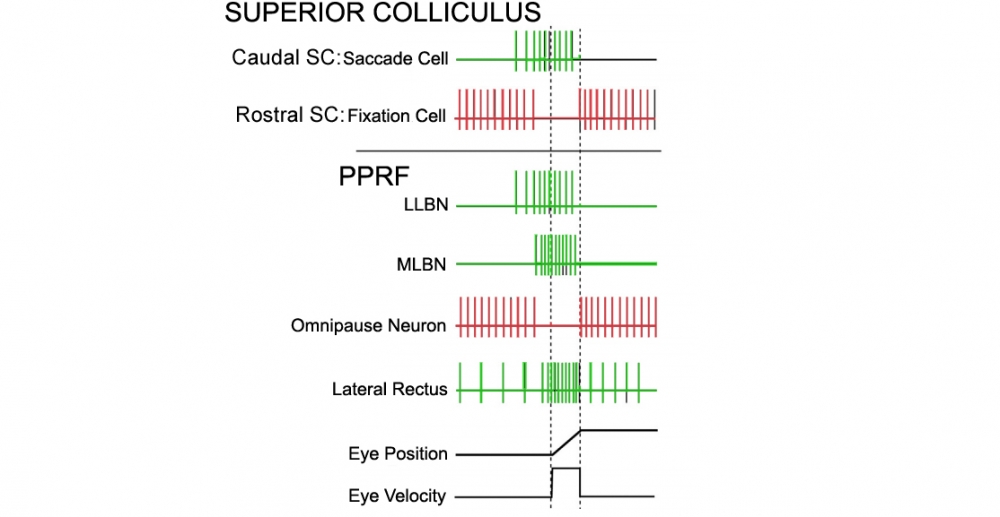
Redrawn from: Shinoda Y, Takahashi M, Sugiuchi Y. Brainstem neural circuits for fixation and generation of saccadic eye movements. Prog Brain Res. 2019;249:95-104. doi:10.1016/bs.pbr.2019.04.007
Overall SC function may be summarized as3 (Figure 2):
- SC identifies the target (in retinotopic coordinates) that is to be foveated
- Generates a trigger signal to shut down the Omnipause Neurons (OPN)
- The SC output neurons for generating saccades activate the brainstem long lead burst neurons (LLBNs), to begin the saccade. LLBNs then sequentiallyl activate the medium lead burst neurons (MLBNs) such as the Excitatory Burst Neuron (EBN)2.
Long Lead Burst Neurons:
These receive inputs from the SC and cortical areas responsible for saccades. LLBN act as a summing junction for combining saccade commands from different areas involved in the preparation for a saccade1. LLBN discharge up to 100 msec before saccade onset.

Medium Lead Burst Neurons: Excitatory and Inhibitory premotor bursts neurons (EBN and IBN)
Two main types of medium lead burst neuron are critical for saccade generation. These are also termed premotor burst neurons (PBN) because they project monosynaptically to ocular motoneurons.
The premotor burst neurons (MLBNs) are:
-The Excitatory PBNs (EBN):
-The Inhibitory burst neurons (IBN):
These premotor burst neurons are silent except during a saccade, when they burst vigorously. MLBNs fire at high frequency 8-15 msec before and during saccades, but are silent during fixation, pursuit, vestibular and optokinetic eye movements5.
Premotor burst neurons are driven by upstream areas, which determine the frequency and duration of the burst, and thus the amplitude, velocity and duration of the saccade. The EBN and IBN have very high gains and can fire at very high frequencies to generate brief bursts, but they can also fire simply by their inhibition having been removed. Note that if there are any delays or noise in the neuronal system, the very high gain of the bursting element would lead to the system tending to oscillate around the target with of a series of overshooting saccades1.
1. The excitatory PBNs (EBN) burst just before saccades and generate the pulse of saccadic innervation. These neurons are glutamatergic.
- Horizontal EBNs are located in the paramedian pontine reticular formation (PPRF), below and extending rostrally from the level of the abducens nucleus. They project monosynaptically to the ipsilateral abducens nucleus, and to a network of neurons in the nucleus prepositus hypoglossi (NPH) and adjacent medial vestibular nucleus (MVN) that contribute to the final common integrator’s horizontal component.
- Vertical and torsional EBNs are located in the midbrain, in the region of the rostral interstitial nucleus of the medial longitudinal fasciculus (RiMLF). They project monosynaptically to the vertical and torsional MNs, and to the interstitial nucleus of Cajal (INC), which is an important part of the final common integrator’s vertical and torsional components. These IBNs are assumed to play a role similar to that of horizontal IBNs, but for vertical and torsional components of saccades. Note that the vertical IBNs are GABAergic, whereas the horizontal IBNs are glycinergic. This difference may contribute to the prevalence of horizontal saccadic oscillations, because glycine is active at two receptors, an inhibitory (strychnine-sensitive) receptor and a neuromodulatory (n-methyl D-aspartate; NMDA) receptor3.
2. The inhibitory burst neurons (IBN):
- Horizontal IBN lie in the medullary reticular formation and extend to the level of the abducens nucleus. The horizontal IBN are glycinergic and project predominantly contralaterally to the abducens nucleus and horizontal EBN, and inhibit the drive to the antagonist muscles.
- Importantly, the IBN also project to the contralateral IBN, which means that the left and right IBN mutually inhibit each other. This reciprocal inhibition can lead to instability and saccadic oscillations4.
- For vertical and torsional saccades, IBN are found in the INC and the riMLF5.
Omnipause neurons (OPN) are an important element of the saccadic system, which inhibit both excitatory and inhibitory burst neurons, resulting in steady fixation of gaze6. LLBN likely inhibit the OPN to release its tonic inhibition on the EBN, thereby allowing the excitatory neurons to discharge.
OPN for horizontal and vertical saccades lie close to the midline in the raphe interpositus nucleus in the PPRF of the caudal pons4. They are glycinergic, and inhibit PBN bilaterally in both the pons and midbrain.
The OPN are active and fire steadily, inhibiting the burst neurons, except during a saccade,and they pause during saccades in any direction. The OPN's function is clearly to prevent the onset of a saccade, but they do not bring about termination of the saccade2. Currently, OPN are believed to act as a neuromodulator to increase the responsiveness of saccade related neurons: they gate the input to the premotor burst neurons. This role is necessary because, without it, neurons in the saccadic system which have very high gains would be susceptible to oscillations at the end of a saccade. By increasing the gain of these neurons only before saccades, OPN can prepare the system to make a fast movement without running the risk of becoming unstable.
It appears likely that lesions of the raphe interpositus region affects omnipause neuron function, resulting in slow saccades, but not involuntary saccades5.
The Frontal Eye Field (FEF) projects both to the superior colliculus and to the brainstem (N. reticularis tegmenti pontis, NRTP). This projects to the cerebellar cortex (oculomotor vermis) (OMV), which has inhibitory projections to the cerebellar fastigial oculomotor nucleus (FOR).
The FOR and SC communicate with omnipause neurons in the pons.
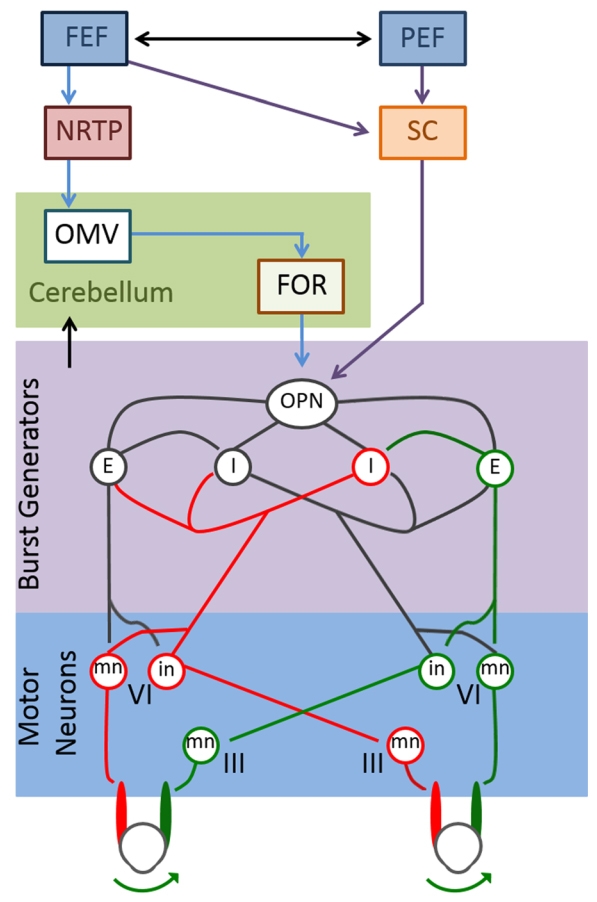
From: Jensen K, Beylergil SB, Shaikh AG. Slow saccades in cerebellar disease. Cerebellum Ataxias. 2019;6:1. Published 2019 Jan 17. doi:10.1186/s40673-018-0095-9