
Smooth-pursuit movements allow clear vision of a moving target by holding the image steady on the fovea. Motion of images across the retina (retinal slip) serves as the primary stimulus for pursuit1, and smooth pursuit evolved with the fovea, making it possible to track small targets that move across a stationary background – something that the optokinetic responses cannot do. Smooth pursuit tracking depends on decoding visual motion, selecting a target of interest and keeping the image of the target on or near the fovea. Motion of the target is critical to pursuit, since attempting to track a static straight line is not possible: this produces a series of saccades2.
Since eye movements themselves cause motion of images of stationary objects on large portions of the retina, an essential component of smooth pursuit is that attention be paid to the moving target relative to the background. Since the pursuit mechanism can distinguish moving foreground objects from the stationary background, this involves the suppression of optokinetic reflexes, since the OKN function is precisely to hold gaze on a stationary background, and prevent the wide-field relative motion which would result from the pursuit of a foreground object3.
Another important property of smooth pursuit is an ability to track targets that are moving with predictable waveforms, such as sine waves; such motion is common in the real world, for example, the swaying of the branch of a tree. Since the latency for smooth pursuit initiation is over 100 ms, perfect tracking implies that the brain must use a prediction mechanism to achieve synchronous tracking of the eye and target1.
Since smooth pursuit allows for tracking of a moving target both when the head is still, and in motion, in addition, the smooth pursuit system needs to cancel the VOR during combined eye-head tracking3: during smooth tracking of a target that moves in the same direction as the head, smooth pursuit cancels VOR; if this were not the case, the VOR would move the eyes in the opposite direction of intended gaze. Deficits in smooth pursuit are usually accompanied by impaired visual cancellation of the VOR.
Pursuit Pathways
The pursuit system has a functional architecture which is similar to that of the saccadic system. Rather than being controlled primarily by areas in extrastriate cortex specialized for processing visual motion, pursuit involves an extended network of cortical areas (eg, frontal eye field) and other subcortical structures (eg, superior colliculus and basal ganglia) that are also important for saccadic eye movements. Thus, although the traditional view is that pursuit and saccades are two distinct systems, it may be more accurate to consider the two movements as different outcomes from a shared cascade of sensorimotor functions3.
With respect to interactions between the saccadic and pursuit systems, a saccade is needed to bring a selected moving target to the fovea, whereafter the pursuit system acts to maintain the target on the fovea4.
Visual information is relayed from the striate cortex to the extrastriate areas (areas V2 and V3), where neurons are specialized for motion, with large receptive fields, strong direction selectivity, and activity that encodes both target and eye motions. From extrastriate cortex, signals are relayed to the area MT (middle temporal, where neurons preferentially respond to the speed and direction of moving stimuli), and then to the medial superior temporal area (MST). The MT and MST areas are part of the dorsal visual processing stream, which plays an important role in detecting moving visual stimuli. Subsequently information is passed to the smooth pursuit regions of the frontal eye field, as well as posterior parietal cortex.

These extrastriate areas have projections via the internal capsulte directly to the brainstem, to the ipsilateral dorsolateral pontine nuclei (DLPN). This, in turn, projects to the cerebellum, to the contralateral flocculus/ventral paraflocculus and the dorsal vermis, underscoring the vital importance of the cerebellum for the generation of smooth pursuit. 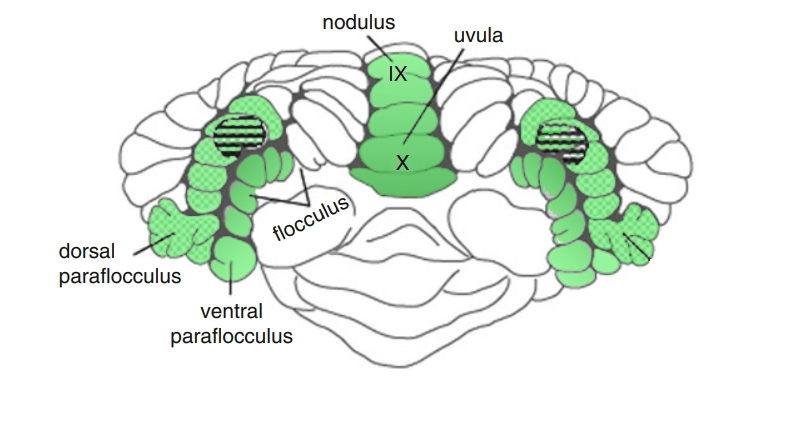
These cerebellar regions project to the medial vestibular nucleus (MVN) and the adjacent nucleus prepositus hypoglossi (NPH), and then to the abducens nucleus and other oculomotor nuclei3.
The final projections from the cerebellum to brainstem structures indicate that the smooth-pursuit pathways within the brainstem and cerebellum share circuitry (MVN and NPH) with the generation of vestibular movements3.
The nucleus prepositus hypoglossi-MVN complex and the INC perform integration, in the mathematical sense, for conjugate horizontal and vertical pursuit, respectively, by transforming eye velocity signals to eye position signals3.
Examination of Pursuit
The patient is asked to track visually an object moving slowly in horizontal and vertical directions (10–200/s) while keeping the head stationary5. The speed of smooth pursuit movements is proportional to target speed. The OKN drum is also a useful method to examine pursuit, in particular to determine if there is any asymmetry present.
Note that most subjects cannot voluntarily generate pursuit movements in the absence of a moving target: the subject needs to be able to fixate the target, in order for normal pursuit to be assessed.
The presence of corrective saccades needs to be determined: these indicate the gain of smooth pursuit (ratio of eye movement velocity to target velocity) as being abnormal:
- Catchup saccdes if gain is too low
- Back-up saccades if gain is if too high.
Smooth pursuit eye movements are most often affected by medication or advancing age, both of which affect the wide range of structures responsible for normal smooth pursuit: visual cortex, medial temporal area, medial superior temporal area, frontal eye fields, dorsolateral pontine nuclei, cerebellum (flocculus), and vestibular and oculomotor nuclei.
VOR cancellation is an additional method to evaluate pursuit.
Pathophysiology of abnormal pursuit
Marked asymmetries of smooth pursuit indicate a structural lesion. For example, if the smooth pursuit is saccadic to the left, this may indicate a left-sided lesion of the flocculus/paraflocculus.
Hemispheric lesions commonly cause pursuit abnormalities. For example, a lesion of area MST gives rise to impaired horizontal pursuit (decreased pursuit gain) towards the side of the lesion (Video)3.
Lesions in different areas of the cerebellum have different effects on smooth pursuit:
- The ventral paraflocculus is the primary structure involved in pursuit and VOR cancellation, and bilateral lesions of the flocculus and ventral paraflocculus cause severe deficits in horizontal and vertical pursuit (in both directions), as well as deficits of VOR cancellation (as seen in spinocerebellar ataxias).
- Lesions of the vermis and fastigial nucleus also lead to deficits in horizontal pursuit.
- Lesions involving the final part of the pursuit pathway (ie, starting from the level of medial vestibular nucleus in the medulla) will also affect the VOR because pursuit and the VOR share similar pathways from this point forward.

Moving the target too fast will compel the patient to use the saccadic system to attempt to follow the target.
(vv)SP2.mp4(tt)
(vv)Smooth_Pursuit.mp4(tt)
(vv)SP.mp4(tt)