

Schematic view of the dorsal aspect of the unfolded cerebellum.

The anterior lobe is separated from the posterior lobe by the primary fissure.
The tonsil sits at the cranio-cervical junction, and the midline is occupied by the cerebellar vermis.
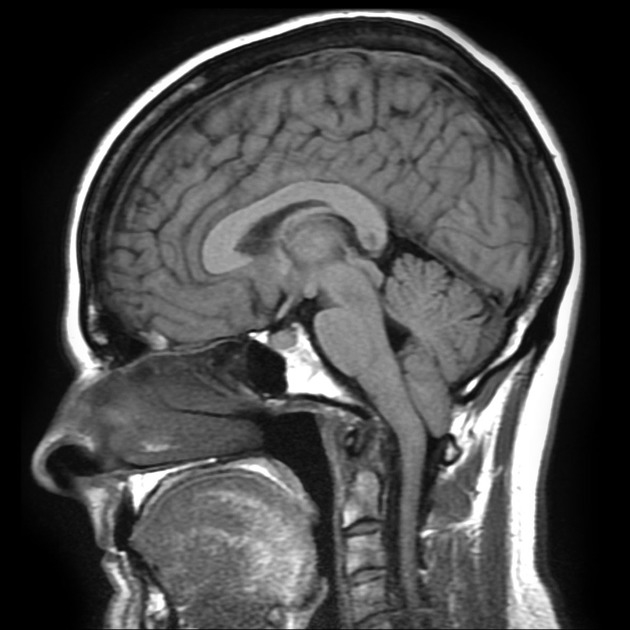
From: Gaillard F. Chiari I malformation. Retrieved from: https://radiopaedia.org/cases/chiari-i-malformation-7
Cerebellar Input
Afferents from motor regions of the cerebral cortex enter the cerebellum via the middle cerebellar peduncle.
Afferents from the spinal cord (spinocerebellar tracts) and from the inferior olive enter the cerebellum via the inferior cerebellar peduncle.
Connections to the cerebellum include primary motor cortex, premotor cortex, and prefrontal cortex. The projections to cerebellum from prefrontal cortex (the apex of the hierarchically organized motor system) are very significant. The cortico-pontine fibres converge in the cerebral peduncle before terminating in the pontine nuclei, which serve as the main relay station for cortical inputs to the cerebellum. Direct projections from cortex, as well as a copy of what is being transmitted via the pyramidal tract, synapse on the pontine nuclei, and then project from the pons to the cerebellum; these are the mossy fibers. This corticopontocerebellar tract forms an extremely dense nerve bundle containing about 20 million axons, about 20 times more than the pyramidal bundle.
The other major input to the cerebellum arises in the inferior olive of the medulla (just lateral to the pyramid). These are the climbing fibres, and pass information to the Purkinje cell.
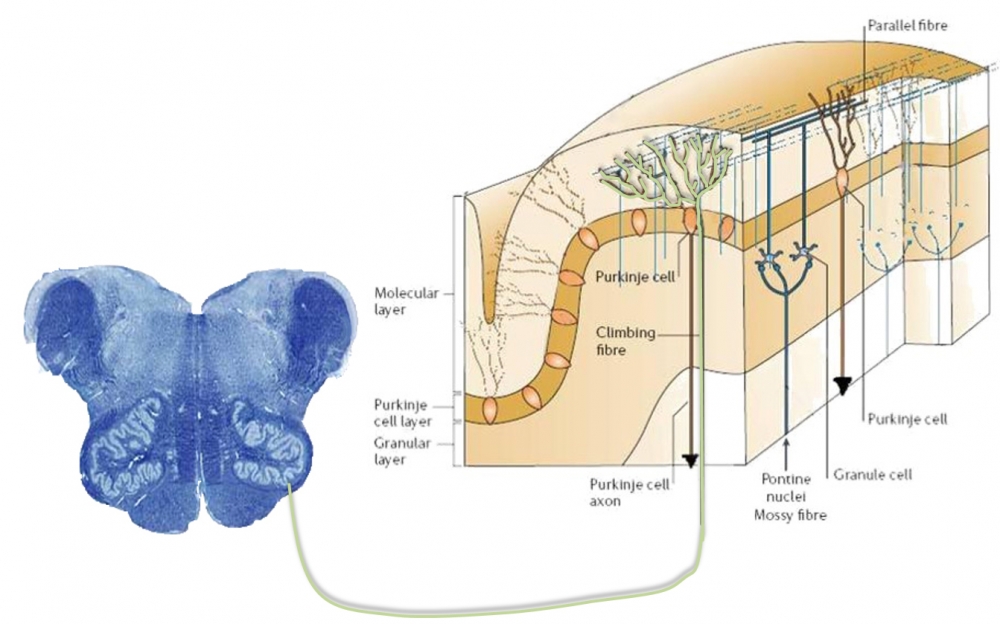
In addition, major sensory input to the cerebellum comes from the spinal cord (spinocerebellar tracts) with updates about the activities of the muscle spindles, tendon organs and joints receptors. This proprioceptive information keeps the cerebellum informed of how current movements have been carried out. Spinal cord afferents particularly project to the vermis and adjacent midline structures.
There is also vestibular nerve input directly to the cerebellum (flocculo-nodular lobe), some of it direct from the nerve, some via the vestibular nuclei (vestibular nuclei can be viewed as ectopic cerebellar nuclei).
Cerebellar Output
Output takes place from the dentate via the superior cerebellar peduncle, crossing the midline and then innervating the “cerebellar” thalamus, and then to motor and premotor cortex.
The deep cerebellar nuclei and vestibular nuclei are the output structures of the cerebellum. Virtually all motor centres of the brain receive excitatory input from the deep nuclei.
- The dentate nuclei project upwards to thalamus and cerebral cortex, and downwards to the spinal cord. The dentate is important in controlling motor activity in the arm and leg.
- The globose and emboliform nuclei send projections to the red nucleus in the midbrain, and give rise to the rubrospinal tract that passes down the spinal cord, adjacent to the corticospinal tract.
- The fastigal nucleus, which receives inhibitory control from the Purkinje cells of the vermis, sends messages to the vestibular nuclei.
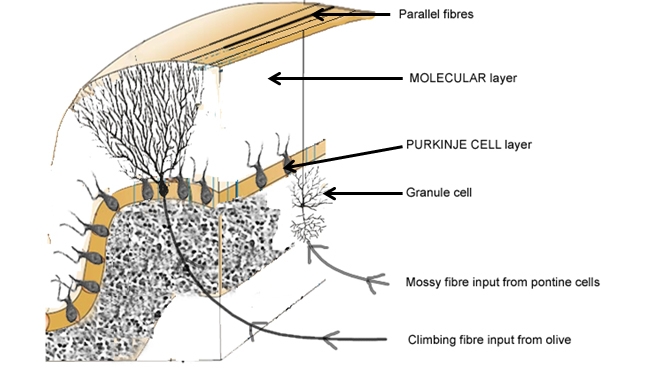
With regard to input to the cerebellar cortex, mossy fibres come from the pontine nuclei and end in a connection with the granule cells. The axons of the granule cells ascend vertically to the molecular layer, where they bifurcate in a ‘T’ junction. The branches on each of the arms of the ‘T’ are known as parallel fibres, and run along the folia to contact the maximal number of Purkinje cells. The parallel fibres drive the Purkinje cells, which have their (inhibitory) output to the deep cerebellar nuclei.
